







Rediscovering Biology: Molecular to Global Perspectives
Online Textbook and Video
This online textbook chapter supports and extends the content of the Genetics of Development video. The chapter examines how the molecules work to coordinate — or misdirect — the formation of an entire organism. Click on the Go button to begin reading, or skip to a sub-section of the chapter in the list below.
1. Genes and Development
“Animals that look nothing like each other develop by using much the same basic ‘toolkit’ of molecules and often in much the same ways.”
– M. Palopoli and N. Patel
Development poses some of the central questions of biology: How does a single cell become a complex multicellular organism like us? What role do our genes play in the processes of development? From the early decades of the twentieth century, geneticists knew about mutants that altered phenotypes because of the actions of various genes during development. In numerous cases biologists knew where on the chromosome the mutant gene was located and how the mutant allele was transmitted from parent to offspring. Nevertheless, the actual role the genes play in development remained a “black box” mystery until around 1980.
Starting in the late 1970s geneticists figured out the details involved in the genetic control of development in model systems such as the fruit fly Drosophila melanogaster. They found that many of these developmental genes shared similar features. During the 1980s and 1990s geneticists made an even more surprising discovery: the same principles, and often the same genes, involved in development in model organisms (such as fruit flies and zebrafish) are also involved in controlling development in most other animals, including humans.
2. Differentiation and Genetic Cascades
Development of a complex multicellular organism is more than just growth — we certainly do not look like gigantic fertilized eggs. Starting from a single cell, numerous specialized cell types emerge that differ in many ways: size, shape, longevity, biochemistry, and so on. What can account for this great diversity among cell types? What processes underlie this differentiation of a single cell into all the cell types of an adult individual?
Is differentiation due to the loss of certain genes in some cell types? While there are some exceptional cases (for example, mature red blood cells lack nuclei), development as a rule is not due to particular cell types having different genes. With only a few exceptions, all the cells in your body contain the same DNA. Discoveries of adult stem cells show that some adult cells retain the potential to produce many, if not all, of the cell types in the organism. These cells can reverse the process of differentiation, reaching a state where their descendants can redifferentiate into all of the cell types.
If cells of an individual are genetically alike, how does differentiation occur? Recall that proteins, not DNA, carry out most cellular functions. (See the Proteins and Proteomics unit.) DNA serves a blueprint from which RNA is transcribed. Proteins come from the amino acid chains that are translated from the RNA. The levels of transcription and translation of a gene determine how much of that gene’s protein will be present in the cell. Gene expression, which encompasses transcription and translation, is the general term to describe the processes in which DNA produces RNA and proteins. It can also include other factors, such as the rate at which RNA is degraded before it can be translated. Differential gene expression will result in varying concentrations and kinds of proteins in cells, causing them to look and function differently. This differential transcription and translation of genes ultimately allows for cellular differentiation. Thus, development is a program that regulates gene expression at the appropriate locations and times.
How is it that, for a given cell type, certain subsets of genes are expressed and other genes are not expressed? As we will see later, the protein product that results from the expression of one gene can influence the expression of several other genes. In turn, the altered expression patterns of these genes can then influence the expression of an even larger number of genes. By this process, called a cascade, a change in one or a few genes can alter the expression patterns of numerous genes.
3. The Details of Gene Expression
What regulates gene expression? The general principles of eukaryotic gene regulation are now well known. Much regulation occurs during transcription as RNA is synthesized from the DNA template. This process is mediated by interactions between proteins and DNA and,
Figure 1. The yellow sphere represents a transcription factor binding to DNA in the nucleus to affect transcription and translation of new proteins.
sometimes, interactions between different proteins. Proteins called transcription factors bind to DNA sequences, known collectively as regulatory elements, located near the coding region of the gene in question
(Fig. 1). When proteins bind to the regulatory elements, it alters the transcriptional machinery and, thus, the level of transcription can change. In some cases the binding of transcription factors to the regulatory elements causes transcription to increase (up-regulation); in other cases it causes transcription to decrease (down-regulation).
The invention of microarray chips in the late 1990s enabled researchers to observe the expression patterns of thousands of genes at the same time. (See the Genomics unit.) Using these chips, researchers can compare the genomic expression patterns of different cell types (such as a neuron versus a liver cell), as well as examine the changes in these patterns that occur as an embryo develops. With the microarray assays, biologists found many previously undiscovered genes that play a role in development. By examining groups of genes that have correlated changes in their expression patterns, biologists have inferred groups of genes that may interact in developmental pathways. They then use other methods to determine
4. Establishing the Gradient and Coordinate Genes
Development is a process where the products of some genes turn other genes on or off. But how does the process start? Even before fertilization, development is occurring. We normally think of an egg as a storehouse of energy supply and nutrients that the embryo will use as it develops. While this is true, the egg also supplies information to establish a molecular coordinate system. This coordinate system provides a way of telling “which end is up”; in other words the location of the embryo’s head is determined even before the egg is fertilized.
Coordinate genes are named because they establish the primary coordinate system for what will become the embryo. One important example of a coordinate gene is bicoid, which is involved in establishing the anterior-posterior polarity in Drosophila. How does bicoid do this? To understand this process we need to first discuss how bicoid gets to the anterior part of the egg.

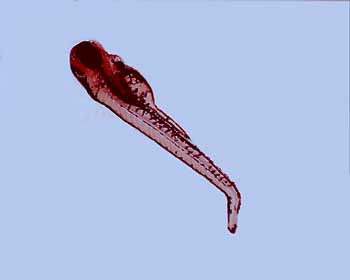
Figure 2. This is a two-hour-old Drosophila embryo that shows the expression of the bicoid protein. The bicoid protein forms a gradient with the highest expression at the anterior end (left side in this photo) of the embryo.
DROSOPHILA EMBRYO WITH BICOID PROTEIN EXPRESSED. Courtesy of Nipam Patel, PhD.
Nurse cells surround the anterior region of the egg in Drosophila and other flies. Cytoplasmic bridges allow various substances – in this case mRNA from bicoid – to be transported from the nurse cells into the egg. The bicoid mRNA is then trapped by proteins produced by other genes. The result is a concentration gradient of bicoid mRNA: the anterior end has the highest concentration and the posterior end lacks it (Fig. 2). Translation of bicoid is inhibited until after fertilization, leading to a bicoid protein concentration gradient.

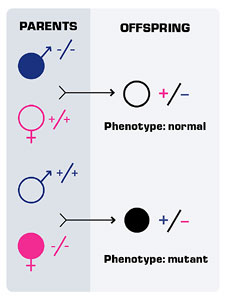
Figure 3. Reciprocal F1 crosses involving maternal effect genes can produce different phenotypes.
In addition to bicoid, other coordinate genes help establish an anterior-posterior polarity. Still, other coordinate genes allow the establishment of a dorsal-ventral gradient. These coordinate genes, like bicoid, are sometimes called maternal effect genes. Maternal effect occurs when the phenotype of the individual is dependent on its mother’s genotype, not its own.
In cases of maternal effect, the transmission pattern of the alleles is the same as in standard Mendelian genetics but the action of the gene occurs a generation later. For example, consider a maternal effect gene where the mutant allele (m) is recessive to the wild-type allele. In the cross of homozygous, wild-type females to homozygous, mutant males, all the F1 offspring are heterozygotes and appear normal. In the reciprocal cross, all of the F1 offspring are heterozygotes but have the mutant phenotype (Fig. 3). Although the F1 offspring are genotypically identical in the reciprocal crosses, they are phenotypically different. This is because phenotype is due to the action of the mother’s genotype. Maternal effect is not the same thing as maternal inheritance, such as in mitochondria, where the genetic material is transmitted only across maternal lines.
5. Responses to the Concentration Gradient
Coordinate genes such as bicoid lay down the grand plan, so to speak, upon which the genes downstream will act. The pattern of the developing embryo arises as these downstream genes are activated or repressed.
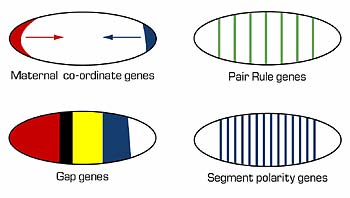
Like many of the other coordinate genes, bicoid encodes a transcription factor; thus, there is a concentration gradient of a transcription factor. The next genes in this developmental cascade, the “gap genes,” possess binding sites for this transcription factor. Gap genes are so named because mutations in these genes can produce larvae with “gaps” (missing several segments). These genes differ in how many bicoid binding sites they have and, thus, vary in their sensitivity to this transcription factor. Some gap genes will become active at

Figure 4. The cascade of developmental genes in segmentation in Drosophila. Maternal effect/coordinate genes set the anterior-posterior axes. The embryo is subdivided into progressively smaller regions by the actions of each class of segmentation genes.
low concentrations of bicoid, while the activation of others will require higher concentrations. Due to the concentration gradient, different regions of the developing embryo will activate different gap genes.
Unlike the coordinate genes, the gap genes are not maternal effect genes. The activities of the embryo’s gap genes (and not those of the mother’s genes) determine the phenotype. Gap genes also encode for transcription factors, and these affect the transcription of genes that further refine the patterning of the Drosophila embryo (Fig. 4).
6. Homeotic Genes
At the end of this cascade is a class of genes that have a long history among Drosophila researchers. Decades before Watson and Crick ascertained the structure of DNA, and even more decades before geneticists understood the principles of gene expression, biologists were using Drosophila melanogaster as a model system for studying the transmission of genetic traits from parent to offspring. Let’s go back to 1915 at Columbia University: In a small laboratory, crowded with thousands of milk bottles containing stocks of the tiny fruitfly Drosophila melanogaster, Thomas Hunt Morgan, the father of Drosophilagenetics, and his students worked. They examined this fruitfly, focusing on ones that looked different in their quest to find and map genes.

Figure 5. When Drosophila has a mutation in the antennapedia gene, legs will grow out of its head.
Thomas Kaufman, PhD, MUTANT DROSOPHILA. Courtesy of Thomas Kaufman, PhD, University of Indiana.
One day Calvin Bridges, one of Morgan’s graduate students, discovered a most unusual fly. One of the hallmark features of flies is that they have two wings; Diptera, the insect order to which flies belong, means “two wings.” The fly Bridges found had one pair of normal wings and one pair of somewhat developed wings. Four wings! Bridges found that this “four wing” phenotype was a genetic mutation which mapped to the third chromosome. After closer inspection, Bridges noted that the third segment of the thorax in these flies looked a good deal like a normal, second segment of the thorax (where wings normally grow). He consequently named the gene associated with this mutant phenotype “bithorax.” (Genes in Drosophila are traditionally named for their mutant phenotype, not for what they do in normal flies.)

Figure 6. Genes that are expressed at the anterior end of an animal are located at the more anterior region of the chromosome. Likewise, posteriorly expressed genes reside on the posterior end of the chromosome. This is referred to as spatial colinearity.
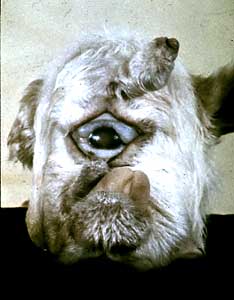
Drosophila geneticists would later find other, similar mutations. One, named ultrabithorax, caused the fly to form two, completely developed pairs of wings. Another, seemingly different mutation (antennapedia), caused legs to grow where the fly’s antennae should have been (Fig.5). These mutant genes became referred to collectively as homeotic genes, named after homeosis. Homeosis, a term coined by William Bateson (a prominent zoologist and one of the early geneticists), refers to “cases in which structures belonging to one body segment were transformed in identity to those belonging to another segment2. Mutants in these genes appeared to change the characteristics of one segment of the fly into those of another segment. Interestingly, all of these genes would map very close together in two clusters on the third chromosome.
Recall the cascade that led to these homeotic genes. The maternal effect coordinate genes laid down the anterior-posterior and dorsal-ventral gradients, which influenced the expression of genes further along the cascade.
These genes turned other genes on or off and, as a result, formed the segmented pattern of the Drosophila embryo. The homeotic genes, having been turned on or off by genes above in the cascade, are also transcription factors. They influence the expression of numerous other genes and, by doing so, determine the identity of the segment they are in. Certain homeotic genes, such as bithorax, are expressed in what would become the thorax; other genes are expressed only in the head or abdomen. It’s interesting to note that genes expressed in similar regions are also located near each other on the chromosome (Fig. 6).
7. Cell Lineage Mapping and C. Elegans
Drosophila melanogaster is not the only model organism for developmental genetic studies. Starting in the 1960s geneticists interested in developmental questions turned to a free-living soil nematode, Caenorhabditis elegans. This species, usually referred to as just C. elegans, has several features that Drosophila and most other organisms don’t have, which makes it attractive for developmental studies. Because embryos of this nematode are transparent, their cells can be observed easily and without much manipulation. The species also has a low number of cells. In fact, all normal individuals have the same number of cells: 959 somatic cells in the hermaphrodite and 1,031 in the male. Unlike Drosophila and mammals, which have extensive cell movement during development, the cells of C. elegans do not move very much during development. All of these features made C. elegans an ideal organism to study cell lineage history, the ancestral-descendant relationship of cells.
John Sulston and colleagues worked out the entire cell lineage history of C. elegans by 1983. Some cell lineage mutations alter the rate and/or timing of cell division. Others affect differentiation. One remarkable feature of C. elegansdevelopment is that seventeen percent of the cells generated during embryogenesis undergo programmed cell death, also called apoptosis. Normal development requires that certain cells die. There are several mutants in which the exact failure of cells to die has been tied to a phenotypic change. Many of the genes involved in programmed cell death in nematodes have counterparts in vertebrates that are also responsible for programmed cell death. Moreover, absence of proper cell death is a key feature of many cancers. (See the Cell Biology and Cancer unit.)
8. Fate Maps

Figure 7a. A photograph of an early stage blastula from the Xenopus laevis frog.
Courtesy of Dr Anna Philpott, Department of Oncology, Cambridge University.
What Sulston and his colleagues did with tracing the entire cell lineage would be exceedingly difficult for the vast majority of organisms. Most multicellular organisms have far more cells than C. elegans. Moreover, most don’t have a transparent body or rather sedentary cells during development. Nevertheless, for several different kinds of organisms, researchers have been able to determine the type of tissue that cells in developing embryos will become; fate maps are diagrammatic representations of this (Fig. 7).

Figure 7b. A representation of a fate map.
Scientists have been able to create fate maps for several organisms (such as the sea urchin) since the early decades of the twentieth century. To construct fate maps researchers use various methods, including removing cells from embryos. If the adult that developed from these embryos is missing specific tissues, researchers infer that the removed cells would have become those missing tissues. Researchers can also use a variety of stains to trace cells in the developing embryo.
9. Cell-Cell Communication and Signal Transduction
Although development begins with a “master plan,” initiated by coordinate genes and carried on through a series of genetic cascades, cells also communicate with one another to coordinate development. In addition, cell-cell communication is essential throughout the life of the organism. Indeed, many cancers are due in part to failures of normal cell-cell communication. (See the Cell Biology and Cancer unit.)
There are some similarities between the way cells communicate and the way individual organisms communicate: in both cases there are signalers and receivers. Cell-cell communication, like many forms of communication between organisms, involves the transfer of information by using molecules between signalers and receivers. The signaling cell sends out molecules called ligands; these can be proteins or small molecules such as vitamin D. Ligands attach to proteins embedded in the membrane of the receiver cell; these proteins are sometimes called receptor proteins.
Once the receptor protein receives the message (the ligand), the nucleus still needs to receive the information because that’s where transcription occurs. How does that happen? Most often, the binding of the ligand causes the receptor protein to change its conformation. This conformational change sets up a series of changes, and sometimes cascades, which eventually lead to changes in transcriptional activity of genes.
One example of a signaling pathway involves the “hedgehog” gene in Drosophila. This gene was so named because larvae with the mutant phenotype are covered with hair and look somewhat like a hedgehog. The protein encoded by the hedgehog gene is a ligand and interacts with several receptors. Among other functions, it triggers the early steps in the development of postsynaptic neurons. It also is involved in the differentiation of the photoreceptor cells of the eye.
10. Conservation of the Homeobox
In the early 1980s Drosophila geneticists started sequencing the DNA from the homeotic genes. Much to their surprise they found that all the homeotic genes contained a 180-basepair region. This region, named the homeobox after the genes in which it was first found, encodes for a sixty-amino-acid sequence that is very well conserved among the homeotic genes. Homeobox refers to the sequence of DNA; the amino acid sequence it encodes is called a homeodomain. Sequences at the homeobox usually differ by ten percent or less between pairs of homeotic genes in Drosophila. Homeoboxes are not restricted to homeotic genes and have been found in several other classes of developmentally important genes. The amino acids encoded by the homeobox region contain a motif called a helix-turn-loop, which is associated with binding to DNA sequences. Thus, a gene with a homeobox would be a prime candidate for a gene that encodes a transcription factor.
More surprising than the discovery of homeoboxes themselves was their ubiquity. Soon after homeoboxes were found in Drosophila, William McGinnis and his colleagues went on a “fishing” expedition looking for homeotic genes. They looked in a variety of organisms using a method called “zoo blotting,” a modified type of Southern blot. (See the Genetically Modified Organisms unit.) The process consisted of using gel electrophoresis to separate the DNA by size from each species they were interested in. The DNA was then heated to separate it into single-stranded DNA (ssDNA). Next the ssDNA was blotted and trapped on nitrocellulose filter paper. The researchers then added single-stranded homeobox DNA, which had been labeled with a radioactive isotope, to the filter paper. If the ssDNA on the blot was sufficiently similar to the labeled homeobox ssDNA, the two ssDNAs would hybridize on the filter paper. The filter paper would be radioactive wherever there was hybridization. To their surprise, McGinnis’s group found homeobox sequences everywhere — in insects, crustaceans, vertebrates (including humans and mice), echinoderms, and mollusks. Almost all multicellular animals had genes with homeoboxes. Moreover, they all expressed these genes during development, often in very similar ways.
Most invertebrates have a single cluster of homeotic genes. In Drosophila that cluster is broken in two. Vertebrates have four copies of the cluster, strongly suggesting that the cluster had been duplicated twice in vertebrates.
11. Conservation of the "Control Switch" Gene for Eyes

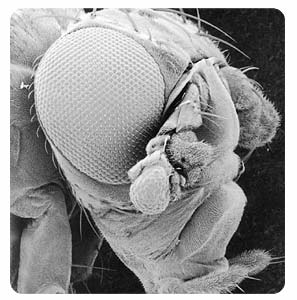
Figure 8. The head of a fruit fly, Drosophila melanogaster, viewed by scanning electron microscope (380x magnification). Targeted expression of the eyeless gene induced the formation of the eye facets on the antenna (to the lower-right of the eye), which are very similar to the facets of the normal eye. This identifies eyeless as the master control gene of eye morphogenesis.
Andreas Hefti and Georg Halder, HEAD OF A FRUIT FLY (1995). Courtesy of Science magazine, cover, 24 March 1995.
Phylogenetic analysis has shown that eyes have independently evolved dozens of times in the history of life. (See the Evolution and Phylogenetics unit.) For example, there are striking differences between the eyes of insects and those of vertebrates. Vertebrates have a camera eye, consisting of a light-sensitive retina, a lens, and a series of muscles used for adjusting focus. In contrast, insects have compound eyes, consisting of numerous light-sensitive ommatidia.
![]()
![]()
Biologists have learned about the genetics of the visual system in insects by studying mutations that affect eyes in Drosophila. Mutants of the eyeless gene in Drosophilahave reduced eye size, with the extent of the reduction depending on the allele. The eyeless gene is normally expressed only in the tissues that become the eyes. Recall that genes in Drosophila are named for the phenotypes of their mutations and not their normal function. What is remarkable about eyeless is that its expression can induce eyes to grow where they ordinarily don’t. Members of Walter Gehring’s lab in Switzerland created transgenic flies which could express the eyeless gene in various places in the developing fly. By expressing eyeless where it is normally not present (ectopic expression), they were able to produce flies with eyes on their antennae, legs, wings, and various other places. So, eyeless looks like a control switch gene for making eyes (Fig. 8).
These same researchers also used databases to search for homologous genes of eyeless in mammals. (See the Genomics unit.) They found that the eyeless gene in Drosophila was strikingly similar (more than ninety percent sequence identity) to the Pax-6 gene in mammals. This Pax-6 gene is also called Smalleyes in mice (where mutants have small eyes) and Aniridia in humans (where mutants lead to deficient development of iris).
Now here’s the really fascinating part! Gehring’s lab did the same ectopic expression experiment but with the mammalian homologue of eyeless. They produced flies with eyes on their antennae, legs, wings, and various other places. The eyes produced were the compound eyes of flies but the machinery for making these eyes could be turned on by mammalian eyeless protein. Despite the independent evolution of eye structure and over
550 million years of independent evolution, the “control switch” for eye development has been conserved.
There are differences between the role eyeless plays in flies and mammals. Unlike in Drosophila, where eyeless is not required for viability, homozygotes for the deletion of eyeless are inviable in mammals. Furthermore, this gene is expressed in regions of the mammalian forebrain. This is strong evidence that eyeless has functions in addition to eye development.
Sonic Hedgehog
Researchers discovered that vertebrates have a homologue to the Drosophila hedgehog gene. They named the vertebrate homologue “Sonic Hedgehog” after the video game character Sonic the Hedgehog. This gene, which encodes a ligand, has diverse functions, including limb development, patterning of the neural tubes (and hence the brain), and differentiation of regions in the gut. How does it work? Cells of the developing notochord send out Sonic Hedgehog signals to the spinal cord. These cells respond to the signaland then differentiate into the ventral part of the spinal cord, which makes the motor neurons that permit muscular activity. Across mammals this gene is highly conserved; the mouse and human Sonic Hedgehog proteins are ninety-two percent identical at the amino acid level.
12. A Brief Look at Plant Development
Despite evolving multicellularity independently, plants and animals share some common features in their respective development. These shared features include homeotic mutations and the use of transcription factors. Research in plant development also started with model organisms — in this case, the mustard grass Arabidopsis thaliana. In Arabidopsis and other plants, the developing flower is composed of four concentric whorls. The outermost whorl (Whorl 1) is fated to become the sepals, the outer floral leaves.

Figure 9. The tissues that will become floral organs are arranged in concentric whorls of a developing flower.
It surrounds Whorl 2, which is fated to become the petals, the white inner floral leaves. Whorl 3 is fated to become stamens, which contains the male organs. The innermost whorl (Whorl 4) is fated to become the carpels, which will form the ovary (Fig. 9).
There are several homeotic mutations in flowers where different parts replace others. For example, in one class of mutations, sepals develop where petals should and carpals develop where stamens should. These mutations have been identified as defects of a family of genes that all encode a particular class of transcription factor, called the MADS box family. MADS box transcription factors occur in both plants and, to a lesser extent, in animals and contain a conserved fifty-eight amino acid sequence.
Plants and animals differ in one important feature: the maintenance of totipotent cells. Cells, like the fertilized egg, which can make all of the cells of the organism, are said to be totipotent. In the process of animal development, the competence of the cells to become different cell types declines. But as cells become more differentiated, they continue to lose competence: In animals, pluripotent cells can produce most, but not all, types of cells, while multipotent cells can produce only a defined set of mature cells. Plants, however, have an apical meristem located at the tip of every root and stem that remains totipotent. They have other meristems that are also totipotent. Moreover, under the right conditions, many differentiated plant cells are able to “de-differentiate to the embryonic state and subsequently redifferentiate to new cell types.”
13. Stem Cells
Certainly some plant cells, like the totipotent meristems, are more versatile than animal cells. Recent discoveries, however, show that the difference in the retention of competence between animals and plant cells is not as great as once thought. During the late 1990s scientists found that adult humans have a reservoir of cells that retain some ability to become other cell types.
Cells derived from fetal tissue have been used to generate so-called embryonic stem cells. In addition to the ethical dilemmas raised by the source of embryonic stem cells, there are practical limitations to the use of these cells for treating and curing diseases and regenerating tissues. Because the donors of these cells are immunologically different from the recipient, immunosuppression would have to be used as in an organ transplantation. Because adult stem cells can be derived from the individual patient, concerns about compatibility of the cells would be obviated.
But do adult stem cells have the same ability to differentiate as embryonic stem cells? Recent studies suggest that adult stem cells may be more versatile than had been previously thought. Catherine Verfaillie and her colleagues at the University Stem Cell Institute derived what they call Multipotent Adult Progenitor Cells (MAPC) from the bone marrow of adult mice. These cells appear to be able to differentiate into virtually all cell types of mouse when injected into mouse blastocysts. These MAPCs have also been injected into living adult mice and have differentiated into liver, lung, and intestine tissue.
14. Coda
The fact that the same principles and many of the same genes direct the development of such different and diverse animals has generated renewed interest and study of how developmental systems evolve. Given the striking similarity of genes used, how do the manifest differences across animals arise and evolve? This question will keep biologists busy for many years to come.
End Notes
-
- Palopoli, M. F. and N. H. Patel. 1996. Neo-Darwinian developmental evolution: can we bridge the gap between pattern and process? Current Opinion in Genetics and Development 6:502-8.
- Raff, R. C. 1996. The Shape of Life: Genes, Development, and the Evolution of Animal Form, 27. Chicago: University of Chicago Press.
- Halder, G., P. Callaerts, and W. J. Gehring. 1995. Induction of ecotopic eyes by targeted expression of the eyeless gene of Drosophila. Science 267:1788-92.