
















Rediscovering Biology: Molecular to Global Perspectives
Online Textbook and Video
This online textbook chapter supports and extends the content of the Biodiversity video. The chapter examines our moral responsibilities to catalog and maintain the world’s ecosystems. Click on the Go button to begin reading, or skip to a sub-section in the list below.
1. Introduction
“It is a somewhat sobering thought that we know more about the number and position of stars in our galaxy, places that none of us will ever visit, than we do about the myriad of small animals that live in our backyard. This is despite the fact that these creatures eat our plants, sometimes bite us but most importantly contribute to the cycling of nutrients that sustain life.”
– Mark Dangerfield 1
Alarmed by the rapid deforestation of the species-rich tropical rain forest, prominent environmental biologists such as the Harvard ecologist Edward Wilson became increasingly active during the 1980s, warning the public about the impending crisis of species loss. In 1986 Wilson and others convened the National Forum on Biodiversity to discuss various problems associated with ecosystem loss. Calling attention to the scope of the crisis, that forum’s organizers coined a new word: biodiversity">biodiversity.
2. What is Biodiversity and Why Should We Conserve It?
The term “biodiversity” was derived from “biological” and “diversity,” and refers to the total diversity of all life in a given locale – one as small as a backyard (or smaller) or as large as the entire planet Earth. One example of a biodiversity measurement is bird watchers listing the species they see in an area on a given day. Although it is often thought of as the number of species in a locale, biodiversity actually has a much wider definition and encompasses levels above and below that of the species. Wilson described biodiversity as the “totality of hereditary variation in life forms, across all levels of organization, from genes to chromosomes within individual species to the array of species themselves and finally at the highest level, the living communities of ecosystems such as forests and lakes.” 2
There is a strong and growing consensus among environmental biologists that we are currently in the midst of a biodiversity crisis. Human-induced global climate change is now accepted as fact. Habitats are rapidly disappearing. Species are going extinct at accelerating rates.
Why should we care about preserving biodiversity? Environmental biologists have outlined two general reasons. First the utilitarian reasons: We rely on a large number of animal, plant, and fungal species for various purposes including food and medicine. In fact, as Simon Levin notes, about forty percent of “all prescription drugs in the United States contain active ingredients originally derived from nature.”3 Moreover, our current knowledge is probably akin to the tip of an iceberg compared to the potential medicinal or other benefits from species that remain undiscovered. This is particularly true with respect to microbes and fungi, which we know less about than plants and animals.
In addition to the benefits from individual species, humans also benefit from maintaining healthy ecosystems; perturbing these ecosystems can adversely affect human health. For instance, Lyme disease emerged in the northeast United States because of changes in the forest ecosystem of that region. As the forests became more fragmented, population sizes of white-footed mice soared because they were now free from competitors or predators, whose populations had declined in the now patchy forests. The mice are a source of blood for ticks, which can carry the Lyme-disease bacterium. As the diversity of other small, ground-dwelling rodents decreased, the mice became an increasingly exclusive source of food for ticks, which also feed on humans and other mammals. This resulted in a surge in the exposure of humans to the bacteria. Through a series of links, forest fragmentation has permitted Lyme disease to rapidly become a major health problem in the eastern United States. (See the Microbial Diversity unit.)
In addition to the utilitarian reasons, there are also non-utilitarian reasons to preserve biodiversity. Part of the beauty of nature comes from the copious diversity of life. Most would agree that a marked reduction in the Earth’s biodiversity would make it a much poorer planet. Related to both the utilitarian and the non-utilitarian reasons is that biodiversity is essentially irreplaceable. The creation of new species by the natural process of speciation usually occurs in time spans of many thousands of generations, far exceeding human lifetimes. The biodiversity that disappears on our watch will be lost not only for our children and their children, but will remain lost for countless generations to follow. In human terms, extinction is forever. Is it moral for humans to cause the irrevocable loss of other species if we can avoid it?
The line between utilitarian and non-utilitarian reasons for preserving biodiversity is blurred. Some reasons now listed as “non-utilitarian” may actually turn out to be utilitarian. Recent research is starting to give us hints that as diversity collapses, the whole ecosystems on which we depend may collapse on a global scale as well. The loss of diversity from a particular area may have a more drastic consequence than simply “it’s not pretty anymore” – it may come to mean, “this is now a wasteland of biological life.”
3. Global Species Diversity
Biodiversity is copious and imperiled, yet it is difficult to measure. This feature makes it also difficult to quantify its loss as well: we know little about what we are losing. Despite its importance, knowledge about biodiversity lags behind that of other areas of science. The statement that opened this chapter echoes those made by several researchers in environmental biology, who have been frustrated by the lack of progress quantifying biodiversity. As we shall see, even the simple question “How many species of animals are on Earth?” has not been answered, even to within an order of magnitude.
Before discussing how scientists address the question “How many species of animals are on the planet Earth?” let’s first ask, “How many species of animals have been described?” There is uncertainty even to the second question’s answer. Some uncertainty reflects differences in opinion among taxonomists about whether different populations are indeed separate species. Some is due to inadequate centralized databases. While there are efforts underway to provide a centralized catalog of described species, none exist as of 2003. The most current estimates are that there are about 1.4 to
1.6 million described species of animals.
What are these 1.6 or so million species of animals? At least one million are insects. A quip from JBS Haldane, polymath and one of the founders of the evolutionary synthesis, illustrates the taxonomic concentration of biodiversity. When asked about what he could divine from nature about the Creator, Haldane replied that he must have had “an inordinate fondness for beetles.” Haldane’s quip was in reference to the sheer quantity of beetle diversity. There are roughly 450,000 different described species of beetles, representing about thirty to forty percent of the known species of insects (Fig. 1). There are about 200,000 described species of flies. In contrast, there are only about 9,000 species of birds and 4,000 species of mammals. Every year, about 2,400 new species of beetles and 1,200 species of flies are described. Thus, the number of species of beetles that scientists will describe in the next five years alone is greater than the total number of current bird species.

Figure 1. A pie chart of the hypothesized distribution of species living on earth today
In addition to animals and plants, biodiversity also includes a vast number of unlabeled species of bacteria, fungi, and protists. These contribute to environmental homeostasis by degrading organic matter and by making the energy in inorganic matter available for growth. Although we often forget these organisms in our consideration of biodiversity, they are critical to the balance and resilience of the environment, especially with respect to their role in nutrient cycles. (See the Microbial Diversity unit.)
Much of the known biodiversity is located in the tropics. In general, species diversity greatly increases as one moves toward the equator: specific hotspots of biodiversity are located in tropical rain forests. Even though they account for only about seven percent of the land area on the planet, tropical rain forests are home to around half the known species of animals.
4. The Erwin Study
Prior to 1982 most biologists thought that the number of undescribed species was roughly comparable to, or perhaps a few times as many as, the number already described. Thus, pre-1982 guesses of the total number of animal species were on the order of several million. But no one really knew.
In 1982 Terry Erwin published a provocative report in which he estimated the number of species of insects to be not several million but an order of magnitude higher – several tens of millions. Erwin reasoned that because the tropical forests appeared to contain vast unexplored areas of biodiversity he would sample there. Erwin, an expert on beetles, fogged the canopy of several trees of the species Luebea seemannii with a pesticide. The fogged insects then fell to the ground, allowing Erwin to sample them. As he sampled the beetles, Erwin kept finding new undescribed species. From the canopy of a single species of tree (L. seemannii) Erwin found more than 1,100 species of beetles.
How did Erwin arrive at a global estimate for the number of species from his “kill ’em and count ’em” experiment? He first estimated that 160 of those species were specialized to the canopy of that particular species of tree. Considering that beetles represent two-fifths of species diversity of insects, there should be about 400 (160 x 5/2) species of insects specialized to the canopy of L. seemannii. This inference assumes that beetle diversity is representative of insect diversity for that species. Erwin assumed that about two-thirds of the insect species were in the canopy and the rest were elsewhere. Based on that assumption, there should be 600 (400 x 3/2) species of insects specialized to L. seemannii. There are an estimated 50,000 species of trees in tropical forests. If each tree has 600 species of insects specialized to it, there should be 30 million species of insects in tropical rain forests.
Many authors expressed criticism and reservations about Erwin’s extrapolations and inferences. Moreover, there have been only a few similar studies, none on the same scale as Erwin’s. Much of the criticism revolves around Erwin’s initial guess that 160 of the species he collected were specialists. If Erwin had overestimated the proportion of specialists, he would be overestimating the total number of species. Likewise, had he underestimated the proportion of specialists, he would have underestimated the total. Nigel Stork noted that Erwin could well be vastly underestimating biodiversity, given that he did not know how much of the diversity of beetles from the L. seemannii he had sampled. Suppose Erwin had only sampled one-third of the beetle diversity; all of his estimates would be three times too low. Could there be 80 million species of animals? 100 million? In actuality, two decades after Erwin’s report, most biologists have revised their estimates for the total number of species downward toward the 10 million range, in part due to studies suggesting that Erwin overestimated the proportion of specialists. Still, nobody really knows how many species are on Earth.
Another factor that adds to the uncertainty about overall global diversity is our lack of knowledge about smaller organisms. There may be hundreds of thousands or millions of mites and fungi that we have literally overlooked. Even less is known about microbes. There are about 5,000 known species of prokaryotes, but scientists estimate that true diversity could range between 400,000 and 4 million species.
5. Seven Kinds of Rarity
Biodiversity is not just the number of species in an area. An area that contained twenty species that were all relatively common would be more diverse than one that contained nineteen rare species and one common species. What do we mean when we say a species is rare? Should it just be based on population size? Deborah Rabinowitz proposed that we should consider rarity along three different axes. The first axis is whether the species has a high or a low population size. The second is whether the species has a large or small geographic range. The third axis is whether the species can occur in a broad range of habitats or whether it is restricted to a more narrow range. According to Rabinowitz, a species could be considered common if, and only if, it had a high population size, large geographic range, and occurred in broad range of habitats; all other species were rare. But they could be rare in different senses. Given that there are three binary criteria, there would be two to the third power, or eight, categories with only one being common; thus, there would be seven different kinds of rarity. Rabinowitz used these criteria to classify wild flower species in Great Britain. While thirty-six percent of the species fell into the “common” category, the most prevalent category comprised species that were widely distributed and had high population sizes, but were restricted in their use of habitat. One lesson from this study is that many species that are abundant and widespread may be subject to extinction if their habitat were degraded.
6. What Factors Determine Extinction Probability?
Other factors being equal, species that have high population sizes are more likely to persist than those with low population sizes. Very small populations are likely to go extinct just by chance in a process called demographic stochasticity. As an extreme case, consider a sexual species that has just two individuals. If both members of the pair are the same gender, it is doomed. Even if the pair does include a male and a female, the species cannot persist unless it produces offspring that are of both genders. The risk of demographic stochasticity leading to extinction is most severe for species with population sizes below about 10 but still is a hazard up until a population size of around 50 to 100, especially for species with low birth rates. Compared with sexual species, demographic stochasticity would be less of a factor for asexuals such as dandelions because a single individual can reproduce without the need for others.
Species with population sizes that number in the hundreds to a few thousand, while not at risk for extinction due to demographic stochasticity, still face other risks. The random evolutionary force of genetic drift reduces genetic variation every generation. The strength of genetic drift is inversely proportional to population size. Thus, species with lower population sizes generally have less genetic variation than their more numerous counterparts. Species that have little genetic variation are at risk of being wiped out by disease. They are also less able to respond to other changes such as global warming. Although there is some disagreement, the consensus is that species with populations above 5,000 are probably safe from extinction because of these genetic factors.

Figure 2. Once a common bird of eastern North America, the last passenger pigeon died in a zoo in 1914.
John J. Audubon (1829). Courtesy of Haley & Steele Art Gallery.
Even species with very large population sizes can go extinct. For instance, a species faces extinction if its habitat is lost and it cannot find a suitable replacement. One striking example is that of the passenger pigeon. During the early 1800s the passenger pigeon (Fig. 2) had a population size in the billions, on the order of the current human population. Overexploitation by hunters and habitat degradation caused its numbers to rapidly dwindle. As its numbers decreased, the species became vulnerable to the genetic factors listed above and then demographic stochasticity. In September 1914, as World War I was beginning, the last passenger pigeon died in captivity. This species went from very abundant to extinct in a century.
7. Keystone Species and the Diversity-Stability Hypothesis
Not all species are equal with respect to their effects on other species. Starfish feeding in the intertidal zone clean an area free of barnacles and mussels. These barnacles and mussels, without predation by the starfish, would come to dominate the community. In a classic 1966 study, Robert Paine removed starfish from enclosures. In those enclosures where the starfish were removed, the number of species in the community dropped from fifteen to eight. Paine called starfish a keystone species">keystone species, one whose presence has a dramatic effect on species diversity.
Prior to 1973 most ecologists thought that more diverse ecosystems would be more stable than would ones with fewer species. This general belief, what has become known as the diversity-stability hypothesis">diversity-stability hypothesis, was based on a variety of observations but not really tested. One such observation was that cultivated land that had simplified ecological communities was more subject to species invasions than similar areas that hadn’t had human influence. In addition, insect outbreaks are much more common in the less diverse boreal forests than they are in tropical forests.
In 1973 Robert May published a theoretical study that challenged the intuitive ideas that ecologists had about the diversity-stability hypothesis. May analyzed randomly constructed communities and found that communities with more species tended to be less, not more, stable. May’s study, like more theoretical studies of the 1970s, assumed that population numbers of each species were at equilibria. This assumption was made, not because it reflected reality, but because it made the mathematics more tractable. More recent studies have shown that if there is some degree of flux in the population numbers, the community can maintain more species than in equilibrium. This variability may allow different species to respond differently to the environment, and can result in fewer species being lost due to competitive exclusion. When theoretical ecologists relax the equilibrium assumption and allow for population fluxes, they have found results consistent with the diversity-stability hypothesis: communities with more species are more stable.
Several lines of evidence now support the diversity-stability hypothesis. The studies conducted by David Tilman and his colleagues provide some of the strongest evidence for the hypothesis. In 1982 Tilman divided grassland fields in Minnesota’s Cedar Creek National History Area into more than 200 plots. He and his colleagues monitored the species richness and community biomass (the total mass of all plants) in each of those fields over the next two decades. They found that diversity within a community is positively correlated with plant community stability, as defined by the extent of variation in community biomass. Various other studies at different scales have found similar results: stability increases with diversity.4
8. Mass Extinctions
Imagine a meteor ten kilometers wide hitting Earth. The resulting impact would cause ferocious tidal waves and massive earthquakes. Sulfuric acid would be released into the air, leading to intensely acidic rain. Later the atmosphere would dramatically cool because of the dust. The impact would affect nearly all life to some extent, and almost certainly there would be a significant decline in biodiversity.

Figure 3. A re-creation by NASA scientists of the impact made by an asteroid 500 miles in diameter at Chicxulub, on the Yucatan Peninsula. This impact is thought to be the cause of the K/T mass extinction 65 million years ago.
RE-CREATION OF CHICXULUB IMPACT (2001). Courtesy of NASA.
Such a scenario is not just the plot of a Hollywood movie like Deep Impact. A meteor that size actually did hit Mexico’s Yucatan peninsula sixty-five million years ago (Fig. 3). The consequences of the impact led to the extinction of many major groups of animals, most notably the dinosaurs. This mass extinction marked the end of the Cretaceous (K) period and the beginning of the Tertiary (T), and is known as the K/T extinction.
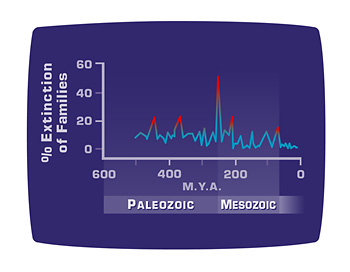
Although the K/T mass extinction is the best known, it was not the largest. That honor belongs to the mass extinction at the end of the Permian period, 250 million years ago. It is often exceedingly difficult to distinguish species in the fossil record, so paleontologists studying extinction usually examine the disappearance of larger taxa (like genera or families). At the end-Permian extinction, sixty percent of families went extinct. Based on the family extinction data, David Raup extrapolated that up to ninety-six percent of species went extinct at this time. Most paleontologists recognize three other mass extinctions, for a total of five (Fig. 4).

Figure 4. Mass extinctions
The graph shows an approximate timeline of loss of families of species from the earth during the five so-called “mass extinctions.”
Although these mass extinctions happened during a short period by geological scales, they were not instantaneous. In fact, the extinctions probably actually occurred over a period of a few million years.
What were the causes of the mass extinctions? We know the most about the asteroid-caused K/T extinction. Based on changes in the floral composition around the K/T boundary, some paleobotanists have speculated that there was global cooling after the extraterrestrial impact. Oceanic cooling may have led to the disappearance of reef-building organisms. We know less about the other extinctions, but it likely that they were marked by periods of global climate change as well.
Species extinctions during mass extinction events account for only a few percent of total extinctions. Indeed, some paleontologists have wondered whether there is anything special about mass extinction events; species extinctions occur often but at different rates across time. Perhaps mass extinctions are merely the tail-end of the distribution of extinction rates.
9. The Sixth Mass Extinction
Should we consider, as some environmental scientists have, that the current biodiversity crisis is the start of a sixth mass extinction? Regardless of how one answers that question, it is clear that we are losing species at rates that, while exceedingly difficult to calculate, are above the background extinction rate and far exceed the speciation rate. Estimates are that 100,000-500,000 species of insects will go extinct in the next 300 years. The higher end of that estimate is comparable to the magnitude of the loss of species during the previous mass extinction episodes. Even the lower estimate represents a considerable loss of biodiversity. Moreover, 300 years is much shorter than the duration of those mass extinction periods.
The current biodiversity crisis stems from several causes. The two major contributors are habitat destruction and global climate change, both of which are largely due to human activity. As discussed earlier, much of the (largely unexplored) biodiversity lies in the tropics and, in particular, tropical rain forests. Tropical forests are being lost at an alarming rate. Conservative estimates place the loss of rain forests during the 1980s and 1990s at about 0.8 percent per year. This is in large part due to changes in the way the land has been used. For quite a long time, many areas had practiced slash and burn agriculture. In recent decades, however, the practice of cutting and clearing has been used increasingly for grazing or timber harvest, resulting in the loss of the tropical forest habitat. As a consequence, countless thousands of species (most of which are unknown to humans) are imperiled.
Global climate change has also impacted biodiversity. During the twentieth century, the mean temperature has increased by slightly more than one degree Fahrenheit (0.6 degree Celsius), and most of that change occurred between 1970 and 2000. Projections vary between x and y degrees Fahrenheit increase by mid-century. These changes do not appear great in the context of daily and seasonal temperature fluctuations, but they are large in comparison with prehistoric climate changes. While the magnitude of these changes is not beyond the range of historical variation, the rate at which the change has taken place appears to be so. The climate change is human-induced, due mainly to increases in carbon dioxide and other “greenhouse gases” that have appeared since the Industrial Revolution and accelerated during the twentieth century.
The human-induced global climate change is coupled with other climate cycles of various temporal and spatial scales. For example, the eastern United States had a cold winter in 2002-3 after several mild winters. In contrast, the western United States had a milder than normal winter that year. The pattern in 2002-3, most likely due to El Niño, does not invalidate the global upward climb in temperatures over a decades-long timespan. In addition to a mean increase in temperature, human-induced global warming is also likely to cause increased variation in climate. Some climate models suggest that the global warming may actually cause the northeastern United States to be cooler. The reason for this seemingly paradoxical possibility is that warming of the oceans could cause the Gulf Stream to be diverted south and east. Were this to happen, it would cause the Atlantic coast to be cooler. Regardless of the specifics of the local changes, more extreme weather will likely exacerbate already fragile ecosystems.
A paper published by Terry Root and her colleagues in 2003 shows that many species have altered their geographic ranges, presumably as a result of global climate change.5 Of those species that had altered their range, eighty percent were in the direction predicted by climate change models. The mean change of movement was about six kilometers per decade. In addition, many bird species have started laying eggs earlier in the spring. This study shows that forces of small, sustained change can be powerful over long enough time scale. But what about species that are unable to move? What will happen as their habitat changes due to human-induced global climate change?
Because of human-induced climate change and habitat destruction, we face a grave and growing crisis. Biodiversity is being lost at alarming but unknown rates. Moreover, if the diversity-stability hypothesis is true, loss of some species may trigger the loss of others, leading to a vicious circle. Although our knowledge about biodiversity and the extent to which it is lost is meager, the consequences are too grave to continue in ignorance.
End Notes
-
- http://www.bio.mq.edu.au/KCBB/AnnualReport/1998.pdf
- Wilson, E. O. 1994. Naturalist. Washington, D. C.: Naturalist Warner Books, 359.
- Levin, S. A. 1999. Fragile dominion: Complexity and the commons. Cambridge, MA: Perseus Books, 6.
- McCann, K. S. 2000. The diversity-stability debate. Nature405:228-33.
- Root, T. L., J. T. Price, K. R. Hall, S. H. Schneider, C. Rosenzweig, and J. A. Pounds. 2003. Fingerprints of global warming on wild animals and plants. Nature 421:57-60.